バナメイエビ(Litopenaeus vannamei)の工場化養殖システムにおける微細藻類群集の特徴の分析(その2)
2017年12月26日
沈 明明: 上海海洋大学水産・生命学院、農業部海洋漁業可持続発展重点実験室 中国水産科学研究院黄海水産研究所
李 健、王 清印、劉 萍、常 志強: 上海海洋大学水産・生命学院、青島海洋科学・技術国家実験室 海洋漁業科学・食物産出過程功能実験室
葛 紅星: 農業部海洋漁業可持続発展重点実験室 中国水産科学研究院黄海水産研究所
( その1よりつづき)
2 結果・分析
2.1 環境理化学的因子の変化状况と養殖状況
養殖期間中の異なる養殖時期の各実験池のpHやアンモニア性窒素などの環境理化学的因子の変化状況は表1に示す通りである。pHの範囲は7.75–8.32で、後期には減少幅が比較的大きかった。全 アンモニア性窒素は0.02–1.79mg/Lで、中期の上昇幅が比較的大きかった。亜硝酸性窒素は0.01–0.71mg/Lで、養殖前期は比較的低い水準にあり、中期に急速に上昇し、後 期は比較的高い水準で安定した。硝酸性窒素は0.01–2.70mg/Lで、養殖の進行に伴い、濃度は比較的急速に上昇した。無機リンは0.01–2.04mg/Lだった。全窒素は1.48–5.65mg/L、全 リンは0.79–4.24mg/Lだった。
| 池番号 | 養殖段階 | pH | アンモニア性窒素 | 硝酸性窒素 | リン酸塩 | 全窒素 | 全リン |
| Pond No. | Culture period | NH+4-N(mg/L) | NO–3-N(mg/L) | PO34–-P(mg/L) | TN(mg/L) | TP(mg/L) | |
| A1 | 前期 Early | 8.24±0.69 | 0.02±0.01 | 0.02±0.01 | 0.04±0.01 | 1.48±0.61 | 0.83±0.24 |
| 中期 Mid | 8.22±0.48 | 0.67±0.25 | 0.68±0.25 | 0.36±0.14 | 2.81±1.46 | 1.39±0.45 | |
| 後期 Late | 7.84±0.62 | 0.41±0.14 | 1.85±0.92 | 1.02±0.61 | 4.86±1.67 | 3.14±1.01 | |
| A2 | 前期 Early | 8.20±0.41 | 0.02±0.01 | 0.02±0.01 | 0.06±0.02 | 1.46±0.63 | 0.93±0.21 |
| 中期 Mid | 8.14±0.62 | 0.68±0.24 | 0.69±0.35 | 0.41±0.17 | 2.72±1.08 | 1.46±0.64 | |
| 後期 Late | 7.82±0.74 | 0.42±0.22 | 1.56±0.42 | 1.25±0.51 | 4.76±1.98 | 3.14±0.93 | |
| B1 | 前期 Early | 7.98±0.61 | 0.03±0.01 | 0.03±0.01 | 0.06±0.02 | 1.64±0.53 | 0.84±0.32 |
| 中期 Mid | 7.88±0.57 | 0.83±0.33 | 0.82±0.34 | 0.56±0.12 | 3.44±1.33 | 1.65±0.65 | |
| 後期 Late | 8.18±0.74 | 0.68±0.28 | 1.93±0.81 | 1.58±0.52 | 4.98±1.09 | 3.82±0.97 | |
| B2 | 前期 Early | 8.22±0.63 | 0.02±0.01 | 0.02±0.01 | 0.04±0.01 | 1.51±0.66 | 0.84±0.23 |
| 中期 Mid | 8.15±0.56 | 0.85±0.34 | 0.91±0.56 | 0.52±0.12 | 2.96±1.26 | 1.09±0.34 | |
| 後期 Late | 7.90±0.86 | 0.56±0.14 | 2.11±1.06 | 1.74±0.65 | 5.21±2.45 | 3.93±1.08 | |
| C1 | 前期 Early | 8.34±0.73 | 0.02±0.01 | 0.02±0.01 | 0.07±0.02 | 1.48±0.69 | 0.79±0.35 |
| 中期 Mid | 8.32±0.76 | 1.78±0.85 | 1.56±0.76 | 0.89±0.32 | 3.48±1.70 | 1.69±0.86 | |
| 後期 Late | 7.75±0.77 | 1.19±0.96 | 2.47±1.02 | 1.98±0.68 | 5.11±2.46 | 4.24±1.38 | |
| C2 | 前期 Early | 8.02±0.85 | 0.03±0.01 | 0.04±0.01 | 0.06±0.01 | 1.61±0.56 | 0.93±0.42 |
| 中期 Mid | 7.98±0.77 | 1.79±1.03 | 1.67±0.64 | 0.78±0.44 | 3.64±1.43 | 1.98±0.58 | |
| 後期 Late | 7.86±0.55 | 1.18±0.75 | 2.45±1.08 | 1.86±0.82 | 5.65±2.53 | 4.03±1.48 |
各養殖池のエビの生存率や生産量などの生長能力は表2に示した通りである。養殖26日目(7月9日)から40日目(7月23日)までは長雨が続き、こ の段階のB1池とC2池のエビは最初の7日で死亡率が40%を超え、その後もエビの死亡が続いた。養殖70日目以後は、B1池とC1池のエビに早期死亡症候群が出現し、養殖が終わるまでエビの死亡が続いた。
| 池番号 Pond No. | A1 | A2 | B1 | B2 | C1 | C2 |
| 生存率 Survival rate (%) | 84.16 | 78.57 | 37.89 | 86.18 | 61.23 | 59.63 |
| 生産量 Yield (kg/m2) | 4.28 | 3.64 | 2.08 | 5.14 | 3.87 | 3.54 |
2.2 微細藻類の種類構成
エビ工場化養殖実験池中の微細藻類の種類と現存量は表3に示した通りである。6つの実験池中には微細藻類5門28属49種が検出された。このうち緑藻植物門は8属10種で総種類数の20.4%を占め、珪 藻植物門は9属19種で38.8%を占め、藍藻植物門は6属13種で26.5%を占め、炎色植物門は4属6種で12.2%を占め、クリプト藻植物門は1属1種で2%を占めた。緑藻と藍藻、珪 藻は多くが優占種と普通種で、炎色植物では2種が優占種、クリプト藻では1種が優占種で、その他は普通種と希少種だった。
| 注:+: 希少種,++: 普通種,+++: 優占種
Notes: +: Rare species, ++: Common species, +++: Dominant species |
|||
| 種類 Species | 現存量 Abundance | 種類 Species | 現存量 Abundance |
| 緑藻植物門 Chlorophyta | C.curvisetus | ++ | |
| プラチモナス属 Platymonas sp. | C. pseudocurvisetus | ++ | |
| P. subcordiformis | ++ | ビドルフィア属 Biddulphia sp. | |
| ユードリナ属 Eudorina sp. | B. sinensis | + | |
| E. elegans | + | B. aurita | + |
| クロレラ属 Chlorella sp. | ディティルム属 Ditylum sp. | ||
| C. vulgaris | +++ | D. brightwellii | + |
| C. pyrenoidesa | +++ | D. sol | + |
| C. ellipsoidea | ++ | フナガタケイソウ属 Navicula sp. | |
| C. kessleri | ++ | N. cryptocephala | + |
| プレオドリナ属 Pleodorina sp. | N. membranacea | + | |
| P. californica | + | 藍藻植物門 Cyanophyta | |
| セレナストルム属 Selenastrum sp. | クロオコックス属 Chroococcus sp. | ||
| S. gracile | + | Ch. minor | + |
| ナンノクロリス属 Nannochloris sp. | Ch. minutus | ++ | |
| N. oculata | +++ | ミクロキスティス属 Microcystis sp. | |
| オオキスティス属 Oocystis sp. | M. marginata | + | |
| O. borgei | + | M. incerta | +++ |
| O. parva | + | M. firma | ++ |
| O. solitavia | + | アファノテーケ属 Aphanothece sp. | |
| コエラストルム属 Coelastrum sp. | Ap. caldariorum | + | |
| C. sphaericum | ++ | Ap. saxicola | + |
| C. microporum | + | ユレモ属 Oscillatoria sp. | |
| ジュズモ属 Chaetomorpha sp. | O. willei | +++ | |
| C. linum | + | O. tenuis | +++ |
| C. crassa | + | O. saline | + |
| ドナリエラ属 Dunaliella sp. | + | O. chlorina | ++ |
| D. salina | + | O. agardhii | + |
| D. bardawil | + | スピルリナ属 Spirulina sp. | |
| 珪藻植物門 Bacillarionphyta | S. major | ++ | |
| メロシラ属 Melosira sp. | フォルミディウム属 Phormidium sp. | ||
| M.sulcata | +++ | P. foveolarum | ++ |
| M.varians | ++ | トリコデスミウム属 Trichodesmium sp. | |
| コアミケイソウ属 Coscinodiscus | T. thiebautii | + | |
| C. lineatus | +++ | 炎色植物門 Pyrrophyta | |
| C. granil | + | プロロセントラム属 Prorocentrum sp. | |
| C. radiatus | + | P. micans | + |
| キクロテラ属 Cyclotella sp. | P. minimum | +++ | |
| C.comta | +++ | ギムノディニウム属 Gymnodinium sp. | |
| C.striata | + | G. coeruleum | + |
| タラシオシラ属 Thalassiosira sp. | G. catenatum | + | |
| T. nordenskioldi | +++ | ペリディニウム属 Peridinium sp. | |
| T. subtilis | ++ | P. depressum | +++ |
| スケレトネマ属 Skeletonema sp. | ケラチウム属 Ceratium sp. | ||
| S.costatum | C. macroceros | ++ | |
| レプトシリンドルス属 Leptocylindrus sp. | C. fucus | ++ | |
| L. minimus | クリプト藻植物門 Cryptophyta | ||
| コレスロン属 Corethron sp. | クリプトモナス属 Cryptomonas sp. | ||
| C. pelagicum | + | C. ovata | +++ |
| キートケロス属 Chaetoceros sp. | クロオモナス属 Chroomonas sp. | ||
| C. lorenzianus | +++ | Ch. oblonga | + |
| C. muelleri | +++ | ||
2.3 微細藻類の数量と多様性の変化
異なる養殖時期のエビ実験池中の微細藻類の密度と生物量、多様度指数は表4に示す通りである。養殖期間全体を通じた6つの実験池の微細藻類の密度は5.2×105–9.4×108cell/L、生 物量は1.23–208.00mg/Lで、微細藻類の密度と生物量はいずれも徐々に高まる傾向を示した。A1池とA2池はエビの密度が最も低く(300ind/m2)、養 殖過程全体の微細藻類の密度と生物量も比較的低い水準にあった。C1池とC2池はエビの密度が最も高く(500ind/m2)、養殖後期の微細藻類の密度と生物量はいずれも比較的高い水準に達し、低 密度池を明らかに上回った。主要種群はユレモで、生物量の占める割合は80%以上に達した。養殖期間の各実験池の微細藻類の多様度指数の範囲は0.42–2.44だった。低密度(A1とA2の間)と中密度( B1とB2の間)の実験池の微細藻類の多様度指数の変化はそれぞれ一定の差異を示したが、2つの高密度(C1とC2の間)の実験池の微細藻類の多様度指数の変化の傾向は一致し、い ずれも養殖時間の経過につれて下降し続けた。
| 池番号
Pond No. |
養殖段階
Culture period |
密度 Density
(106 cell/L) |
生物量
Biomass (mg/L) |
多様度指数 H' |
| A1 | 前期 Early | 2.31±0.53 | 6.92±2.63 | 1.56±0.24 |
| 中期 Mid | 13.89±3.86 | 8.43±1.62 | 0.75±0.05 | |
| 後期 Late | 96.63±24.87 | 134.32±119.23 | 1.28±0.45 | |
| A2 | 前期 Early | 0.64±0.13 | 1.23±0.27 | 0.68±0.07 |
| 中期 Mid | 4.32±1.76 | 4.76±2.13 | 0.95±0.18 | |
| 後期 Late | 12.65±5.12 | 10.80±6.27 | 0.74±0.26 | |
| B1 | 前期 Early | 2.16±0.39 | 1.04±0.33 | 1.59±0.39 |
| 中期 Mid | 121.28±58.90 | 38.34±19.23 | 0.48±0.00 | |
| 後期 Late | 562.23±276.97 | 168.65±58.23 | 1.06±0.29 | |
| B2 | 前期 Early | 1.89±0.57 | 5.23±2.76 | 1.94±0.28 |
| 中期 Mid | 28.64±13.34 | 34.20±15.21 | 2.35±1.22 | |
| 後期 Late | 86.86±58.22 | 136.80±119.83 | 1.32±0.42 | |
| C1 | 前期 Early | 3.41±1.25 | 8.95±6.83 | 1.65±0.35 |
| 中期 Mid | 476.65±128.23 | 75.64±47.32 | 1.49±0.62 | |
| 後期 Late | 869.32±493.73 | 186.56±154.98 | 0.79±0.13 | |
| C2 | 前期 Early | 2.75±1.23 | 6.84±5.23 | 1.13±0.36 |
| 中期 Mid | 133.48±123.27 | 45.57±23.85 | 0.85±0.05 | |
| 後期 Late | 652.32±286.22 | 153.40±95.83 | 0.63±0.06 |
2.4 エビ養殖池の微細藻類の優占種の構成と遷移状況
異なる養殖時期の各エビ養殖池の微細藻類の優占種の遷移状況は図1に示す通りである。優占種とそれに対応するコードは表5に示した。
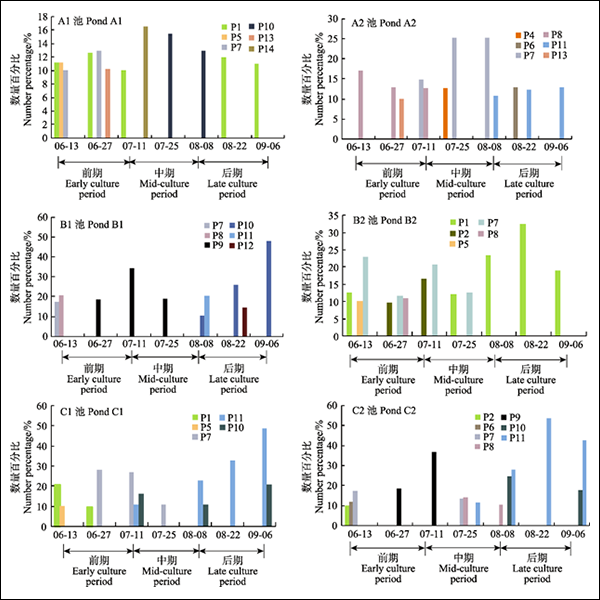
図1 エビ養殖池の微細藻類の優占種の動的遷移
Fig.1 Dynamic succession of microalgae species in shrimp ponds
P1–P14は微細藻類の優占種のコードを示す。品種とコードの対応は表5の通りである。
P1–P14 in this diagram represented corresponding codes for dominant species, and microalgae species, and their corresponding codes were shown in Tab.5
| コード
No. |
微細藻類の種類 Species |
学名 Latin names |
コード
No. |
微細藻類の種類 Species |
学名 Latin names |
| P1 | クロレラ・
ブルガリス |
Chlorella vulgaris | P8 | キートケロス・
ムエレリ |
Chaetoceros muelleri |
| P2 | クロレラ・
ピレノイドサ |
Chlorella pyrenoidesa | P9 | ミクロキスティス・
インケルタ |
Microcystis incerta |
| P3 | ナンノクロリス・
オクラタ |
Nannochloris oculata | P10 | オスキラトリア・
ウィレイ |
Oscillatoria willei |
| P4 | コスキノディスクス・ リネアトゥス |
Coscinodiscus lineatus | P11 | オスキラトリア・
テヌイス |
Oscillatoria tenuis |
| P5 | キクロテラ・ コムタ |
Cyclotella comta | P12 | プロロセントラム・
ミニマム |
Prorocentrum minimum |
| P6 | タラシオシラ・ ノルデンスキオルディー |
Thalassiosira nordenskioldi | P13 | ペリディニウム・
デプレッサム |
Peridinium depressum |
| P7 | キートケロス・
ロレンツィアヌス |
Chaetoceros lorenzianus | P14 | クリプトモナス・
オバタ |
Cryptomonas ovata |
低密度の養殖実験池A1池の優占種は6種だった。養殖前期には、クロレラ・ブルガリスが優勢を占め、このほかキクロテラ・コムタ(C. comta)とペリディニウム・デプレッサム(P. depressum)、クリプトモナス・オバタ(C. ovata)が見られた。優占種の優占度はいずれも0. 15以下だった。養殖42日目となると、藍藻植物門のオスキラトリア・ウィレイ( Oscillatoria willei)が優占種となったが優占度は次第に下がり、クロレラ・ブルガリスが再び優占種となり、養殖末期まで優占種であり続けた。A2池の優占種は6種だった。養 殖段階全体を通じて微細藻類の密度はほかの池を明らかに下回った。前中期の優占種は主に、珪藻植物門のコスキノディスクス・リネアトゥス(C. lineatus)とキートケロス・ムエレリ(C. muelleri)だった。14日目にはペリディニウム・デプレッサムとキートケロス・ムエレリがともに優占種となった。後期は、藍藻植物門のオスキラトリア・テヌイス(O. tenuis)が 長期にわたって優勢を占めた。
中密度の養殖実験池B1池の優占種は6種だった。前期は、キートケロス・ムエレリが主な優占種となった。14日目以後は、ミクロキスティス・インケルタ(M. incerta)が急速に優占種になり、こ の状況が50日目まで続いた。その後、優占種はオスキラトリア・ウィレイとオスキラトリア・テヌイスに変わった。プロロセントラム・ミニマム(P. minimum)は 養殖70日目前後に優占種となったがこれは5日しか続かなかった。オスキラトリア・ウィレイは優占度が養殖末期に0.48に達し、絶対優占種となった。B2池の優占種は5種だった。前中期の優占種は主に、緑 藻植物門のクロレラ・ブルガリスと珪藻植物門のキクロテラ・コムタとキートケロス・ロレンツィアヌス(C. lorenzianus)だった。後期はクロレラ・ブルガリスが絶対優占種となり、優 占度は0.32に達した。
高密度の養殖実験池C1池の優占種は5種だった。前期の優占種はクロレラ・ブルガリスとキクロテラ・コムタ、キートケロス・ロレンツィアヌスだった。28日目前後には、藍藻植物門のオスキラトリア・テ ヌイスとオスキラトリア・ウィレイが徐々に優占種となり、養殖後期には絶対優占種となった。オスキラトリアの優占度は0.82に達した。C2池の優占種は7種だった。養殖14日目までの優占種はクロレラ・ピ レノイドサ(C. pyrenoidesa)とタラシオシラ・ノルデンスキオルディー(T. nordenskioldi)、キートケロス・ムエレリだった。その後、ミクロキスティス・イ ンケルタが急速に絶対優占種となった。28日目には、珪藻植物門のキートケロスとオスキラトリア・テヌイスが優占種となった。38日目には、オスキラトリアがキートケロスに代わって絶対優占種となった。
3 検討
本研究では、各実験池中の微細藻類の数量の変化はいずれも、前期には比較的低く、中期には急速に高まり、後期に最高に達するという傾向を示した。青島地区のバナメイエビ工場化養殖では、地 下海水を砂ろ過して用いている。養殖池に入る前にはさらにスクリーンろ過がなされる。多層的なろ過を経た水は微細藻類の含有量が少ない。このため養殖前期の密度は低くなる。養殖中後期には水温が徐々に高まる。さ らにエビの急速な生長と投餌量の増加に伴い、摂食されなかった大量の残料やエビの排泄物などの栄養塩が養殖池に累積され、微細藻類の大量の繁殖を刺激する(劉孝竹ら,2011; 彭聡聡ら,2011)。C 1池で最後に採取したサンプルでは、ユレモ(オスキラトリア)の生物量が165mg/Lに達し、微細藻類の総生物量の78%を占めた。C2池のユレモの生物量も119mg/Lに達し、63%を占めた。水中には、ユ レモの糸状の群体が塊となって壁に張り付いて生長しているのが見られ、大量のエビが発病・死亡した。拠曹平ら(2011)の報告によると、ユレモの濃度が12.5mg/Lに達すると、エ ビの生存率は際立って低下する。ユレモもアンモニア性窒素や亜硝酸性窒素などの有害物質を吸収できるが、高密度のユレモは「倒藻」(藻類が死亡し水が濁る現象)を引き起こす可能性が高く、ヒ ドロキシルアミンや硫化物などの有毒物質が大量に放出され、エビのストレス反応による死亡を引き起こす(Cremenetal,2007)。このため養殖後期には、ユレモなどの不良な微細藻類に注意し、繁 殖の評価と有効な制御を行う必要がある。A2池の微細藻類の生物量はほかの養殖池より明らかに低い。エビは発病しなくても生長が遅く、生産量は明らかに同程度の養殖密度のA1池より低い。原因は、低 密度の藻類相構造によって養殖システムに対する微細藻類の調節能力が低下し、水質が変わりやすく不安定となり、エビの生長に不利な影響がもたらされるためと考えられる。
多様度指数は、浮游植物の異質性と安定性を定量的に反映するのに用いることができる。指数が大きいほど、微細藻類の群集の情報量が大きく、干渉抵抗能力が強く、安定性が高い(孫耀ら,1998)。中 高密度のエビ実験池の微細藻類の多様度指数は、最初は高まって後に低下するという傾向を示した。これは、養殖前期は水体が多層的なろ過を経ていることから、池中の微細藻類の種類と数量が極めて少なく、多 様度指数も低いが、養殖の進行に伴い、生態系が少しずつ改善し、安定性が増強し、多様度指数もこれに応じて増加するためと考えられる。養殖後期となると、藍藻類のユレモが暴発的に生長して優占種となり、微 細藻類群集は単一方向へと遷移し、多様度指数は急速に低下する(彭聡聡ら,2011)。B1池の中後期とC1池・C2池の後期では、多様度指数がほぼ0.9を下回り、水質がさまざまな程度悪化し、エ ビの発症や死亡の現象が出現する。その他のエビ養殖池では、何回か0.9を下回ったのを除けば、ほぼ0.9以上で、エビの生長は良好だった。これは郭皓ら(1996)の報告の結論と相似している。こ のため養殖過程においては、微細藻類の多様度指数が0.9を下回った際には、管理を強化し、エビの発病を予防する必要がある。
優占種は、一つの群集の生態の特徴を代表する(黄翔鵠ら,2002)。本研究では、エビ工場化養殖の中後期に、優占種となったユレモが総量の58%–90%を占めた。微細藻類の総量の変動は、比 較的単一な優占種によって制御されることを示している。張才学ら(2007)の調査によると、湛江地区のエビ工場化養殖池の後期では、微細藻類の優占種は主に、クロレラやキクロテラなどの耐汚染性の品種だった。李 玉全ら(2006)が青島小麦島で行った研究では、エビ工場化養殖池中の微細藻類群集は珪藻が主だった。本研究では、前期の優占種は主に、緑藻と珪藻の品種が中心で、中後期は多くが、藍 藻のミクロキスティスとユレモが優占種となり、以上の二者の研究結果とは大きく異なった。これは、後期の水体の高栄養水準がユレモなどの藍藻の大量発生の原因となり、藍 藻の大量繁殖によって緑藻や珪藻などの生長が抑えられたためと考えられる(彭聡聡ら,2010)。エビ工場化養殖モデルは、小規模な水中において行われる高密度な商業化養殖の方式であり、その養殖過程は、投 餌や水交換、酸素投与、消毒などのさまざまな人工的因子に左右される(孟慶武ら,2008)。養殖地区の違いや養殖技術や養殖用水の処理方式の違いなども、微細藻類の優占種の差異をもたらし得る(呉斌ら,2 008; 李雪松ら,2006)。
本研究では、微細藻類の群集の遷移に対して養殖密度も大きく影響することがわかった。比較的低い養殖密度のA1池(300ind/m2)では、ほとんどの期間、クロレラ・ブルガリスが優勢となり、藻 類相は良好だった。だがB1池(400ind/m2)とC1池・C2池(500ind/m2)は養殖後期、藻類相がユレモによって支配された。盧静ら(2000)の報告によると、低密度(70ind/m2)の 養殖囲いでは、微細藻類は、珪藻の仲間であるフナガタケイソウ(Navicule)とキクロテラが中心となり、中高養殖密度(210–350ind/m2)のエビ養殖池では、ギムノディニウム属( Gymnodinium)やクロオコックス属(Chroococcus)などの品種が優勢となる。李玉全ら(2013)の報告によると、エビ養殖密度は、工場化養殖システムの窒素・リ ンのアウトプットに大きく影響する。養殖密度が高まると、投餌量やエビの代謝物や排泄物がいずれも低養殖密度のものよりも高くなり、高密度のエビ養殖池の栄養水準は低密度のエビ養殖池よりも高くなり、水 中の栄養水準の違いは微細藻類の遷移の差異を招くこととなる。以上のように、高すぎる養殖密度は、養殖後期に藍藻などの不良な微細藻類が優占種に遷移する原因の一つとなる可能性がある。
(おわり)
参考文献:
Attayde JL, Hansson LA. Effects of nutrient recycling by zooplankton and fish on phytoplankton communities. Oecologia, 1999, 121(1): 47–54
Cai LT, Chen C, Wang YN, et al. Dynamic change of the phytoplankton in the breeding ponds of Penaeus vannamei. Journal of Ningbo University (Natural Science and Engineering Edition), 2013, 26(2): 7–11 [蔡林婷・陳晨・王一農等. 凡納濱対蝦設施化養殖塘中浮游植物的動態変化. 寧波大学学報(理工版), 2013, 26(2): 7–11]
Cao P, Huang XH, Li CL, et al. Effects of Oscillatoria sp. on growth and immune activities of Litopeuaeus vauuamei. Fishery Modernization, 2011(5): 25–30 [曹平・黄翔鵠・李長玲等. 顫藻対凡納濱対蝦生長和免疫相関酶活力的影響. 漁業現代化, 2011(5): 25–30]
Chen JF, Lai YH, Tong WP. Fluctuation of pH values in water of industrialized cultivation of white leg shrimp Litopenaeus vannamei. Fisheries Science, 2006, 25(9): 457–458 [陳剣鋒・頼廷和・童万平. 凡納濱対蝦工厰化養殖水体pH値的変化特徴. 水産科学, 2006, 25(9): 457–458]
Cremen MC, Martinez-Goss MR, Corre VL, et al. Phytoplankton bloom in commercial shrimp ponds using green-water technology. Journal of Applied Phycology, 2007, 19(6): 615–624
Guan CW, Liu H, Zhang YL. Experimental study on breeding Litopenaeus vannamei in recirculating aquaculture system. Fishery Modernization, 2010, 38(4): 21–25 [管崇武・劉晃・張 宇雷. 凡納濱対蝦工厰化循環水養殖試験研究. 漁業現代化, 2010, 38(4): 21–25]
Guo H, Yu ZG. The characterisitics of phytoplankton community in prawn pond and the relationships with shrimp disease. Marine Science, 1996(1): 39–45 [郭皓・于占国. 蝦池浮游植物群落特徴及其與蝦病的関係. 海洋科学, 1996(1): 39–45]
Guo YJ. Chinese algae. Beijing: Science Press, 2003 [郭玉潔. 中国海藻志. 北京: 科学出版社, 2003]
Huang XH, Wang QH. A study on dominant phytoplankton species in high-level prawn ponds and its formation cause. Journal of Tropical Oceanography, 2002(4): 36–44 [黄翔鵠・王慶恒. 対蝦高位池優勢浮游植物種群與成因研究. 熱帯海洋学報, 2002(4): 36–44]
Li XS, Liang JR, Chen CP, et al. Species diversity of phytoplanktons in shrimp aquaculture areas in the Quanzhou Bay Fujian Province. Journal of Xiamen University (Natural Science), 2006(S1): 234–239 [李雪松・梁君栄・陳長平等. 泉州湾蝦池浮游植物種類多様性研究. 厦門大学学報(自然科学版), 2006(S1): 234–2 39]
Li YQ, Li J, Wang QY, et al. Analysis on structure and benefit of super-intensive and semi-intensive shrimp farming systems. Marine Fisheries Research, 2006, 27(5): 85–90 [李玉全・李健・王清印等. 対蝦工厰化養殖與池塘養殖系統結構與効益比較分析. 海洋水産研究, 2006, 27(5): 85–90]
Li YQ, Wang RJ, Jiang LX. Effects of density-stress on water quality and growth of Marsupenaeus japonicas. Marine Fisheries Research, 2013, 37(10): 53–57 [李玉全・王仁傑・姜令緒. 密度脅迫対日本嚢対蝦生長和水環境的影響. 海洋科学, 2013, 37(10): 53–57]
Liu J, Qu KM, Liu HY, et al. Studies on features of water environment in industrialized shrimp culture system. Marine Fisheries Research, 2008, 29(6): 1–8 [劉嬌・曲克明・劉海英ら. 対蝦工厰化養殖水環境基本特徴研究. 海洋水産研究, 2008, 29(6): 1–8]
Liu XZ, Cao YC, Li ZJ, et al. Study on microalgae community structure and its relationship with environment factors in the shrimp ponds during the later phases of culture period. Progress in Fishery Sciences, 2011, 32(3): 84–91 [劉孝竹, 曹煜成, 李卓佳等. 高位蝦池養殖後期浮游微藻群落結構特徴. 漁業科学進展, 2011, 32(3): 84–91]
Lu J, Li DS, Dong SL. The impact of stocking density on plankton community in shrimp pond. Journal of Fisheries of China, 2000, 24(3): 240–246 [盧静・李徳尚・董双林. 対蝦池的放養密度対浮游生物群落的影響. 水産学報, 2000, 24(3): 240–246]
Meng QW, Wang S, Zhao YJ, et al. The characteristics, development trend and prospects of prawn's industrial aquaculture in China. Ocean Development and Management, 2008, 25(3): 124–127 [孟慶武・王聖・趙玉潔等. 我国対蝦工厰化養殖的特点、発展趨勢及前景. 海洋開発與管理, 2008, 25(3): 124–127]
Peng CC, Li ZJ, Cao YC, et al. Change of dominant species of planktonic microalgae in Litopenaeus vannamei's semi- intensive culture ponds and its impact on the culture environment. Marine Environmental Science, 2011, 30(2): 193–198 [彭聡聡・李卓佳・曹煜成ら. 凡納濱対蝦半集約化養殖池塘微藻優勢種変動規律及其対養殖環境的影響. 海洋環境科学, 2011, 30(2): 193–198]
Sun Y, Li F, Li J, et al. Phytoplankton community characteristics and the interrelationship with nutrient salts in shrimp ponds. Marine Fisheries Research, 1998, 19(2): 45–50 [孫耀・李鋒・李健等. 蝦塘水体中浮游植物群落特徴及其與営養状况的関係. 海洋水産研究, 1998, 19(2): 45–51]
Wu B, Liao SM. Primary report of species composition of microalgae in the culture ponds of shrimp Litopenaeus vannamei in Beihai, Guangxi. Guangxi Sciences, 2008, 15(4): 452–455 [呉斌・廖思明. 広西北海凡納濱対蝦養殖池塘中微型藻類組成調査. 広西科学, 2008, 15(4): 452–455]
Xie LM, Lin XT, Xu ZN, et al. Phytoplankton communities and physical and chemical factors in different types of shrimp ponds. Ecologic Science, 2003, 22(1): 34–37 [謝立民・林 小涛・許忠能ら. 不同類型蝦池的理化因子及浮游植物群落的調査. 生態科学, 2003, 22(1): 34–37]
Zhang CX, Lao Z, Liu YL, et al. Change of phytoplankton and physicochemical factors in ponds of shrimp Peuaeus vauuamei with different cultural patterns during late period. Journal of Guangdong Ocean University, 2007, 27(4): 38–44 [張才学・労賛・劉玉蓮等. 凡納濱対蝦常見養殖模式下養殖後期浮游植物及理化因子的変化. 広東海洋 大学学報, 2007, 27(4): 38–44]
Zhao W. Hydrobiology. Beijing: China Agriculture Press, 2005 [趙文. 水生生物学. 北京: 中国農業出版社, 2005]
※本稿は沈明明, 李健, 王清印, 葛紅星, 劉萍, 常志強「凡納濱対蝦(Litopenaeus vannamei)工廠化養殖系統微藻的群落特征分析」(『漁業科学進展』2 017年第38卷第5期、pp.64-72)を『漁業科学進展』編集部の許可を得て日本語訳・転載したものである。記事提供:同方知網(北京)技術有限公司